Les merveilles de la procréation humaine

Le tableau ci-dessous représente la progression et les transformations successives que va subir l’ovule durant la période pendant laquelle il va rester en suspension dans l’organisme maternel (ovocyte 1 au moment de l’ovulation avec 23 chromosomes, puis ovocyte 2 après fécondation et 2 fois 23 chromosomes, il est appelé cellule zygote après fusion des données parentales avec 46 chromosomes). Il va cheminer dans la trompe de Fallope jusqu’à l’endomètre de l’utérus (muqueuses utérines) dans lequel il s’implantera (nidation). Durant toute cette période, il pourrait donc être évacué par une forme de balayage puisqu’il n’est en rien solidaire du corps de la femme. Tout est ainsi mis en œuvre contre toute forme d’agression extérieure de cet ovule, en le conservant à l’intérieur du corps de la femme, mais également contre toute auto-défense immunitaire de la mère à l’encontre des spermatozoïdes puis de la nouvelle cellule qui sont tous deux d’une génétique différente de celle du corps de la femme.
En ce qui concerne la protection contre le système immunitaire maternel du zygote et ses 46 chromosomes, dont 23 sont différents, celui-ci reste entouré d’une peau appelée zone pellucide issue de l’ovule et des cellules péri-ovocytaires issues du follicule, qui contiennent au minimum 23 chromosomes maternels, voir les 46 pour les cellules péri-ovocytaires. La zone pellucide est particulièrement conçue pour assurer un rôle protecteur contre la polyspermie (double introduction de spermatozoïde humain), ainsi que toute intrusion d’un spermatozoïde étranger. Contre la polyspermie, l’ovule dispose d’une première barrière constituée par le tissu cellulaire perméable des cellules péri-ovocytaires, qui vont limiter le nombre de contacts simultanés des spermatozoïdes, la seconde est la membrane plasmique rendue infranchissable après une première introduction.
Si la zone pellucide s’avère donc être un très bon système protecteur et sélectif durant la phase où l’ensemble embryonnaire se trouve en suspension dans le corps de la femme, elle ne peut être cependant que temporaire, puisqu’elle ne permettrait pas l’implantation de tout le système nourricier nécessaire à la survie et la croissance de l’embryon. C’est pourquoi, durant cette phase « volatile », la division du zygote (segmentation) avec ses 46 chromosomes va lui permettre de se fabriquer une protection compatible tant avec le système immunitaire de la mère qu’avec le sien.
Comme nous pouvons le voir en effet au niveau de la dernière phase avant implantation, ce n’est que lorsque le zygote se sera divisée suffisamment et produit des cellules spécialisées pour générer une poche protectrice, future placenta, qu’il pourra-t-être implanté à nouveau dans l’organisme féminin sans en être rejeté. Cette durée est bien évidemment variable en fonction de la femme concernée, tout comme la division du zygote est variable.
Ce qui est remarquable à notre niveau, c’est de percevoir combien toute cette logique de conception est déjà inscrite jusqu’au moindre détail dans cette première cellule lorsqu’elle est fécondée, puisque c’est à partir de celle-ci qui mesure seulement un dixième de millimètre que pourra se reproduire chez le nouvel individu à l’état adulte tout ce même processus s’il est de nature femelle. Un laps de temps important se sera passé entre l’état d’ovule et celui de femme adulte, mais la logique inscrite dans l’ovocyte 1 sera restée la même afin de générer un individu capable de procréer à son tour.
6 – Évolution de l’ovule fécondé
L’ovule, également appelé ovocyte 1 à ce stade, est présent dans chaque follicule et grandit avec celui-ci à raison d’un par cycle (sauf faux jumeaux), depuis la puberté jusqu’à la ménopause de la femme. Après l’ovulation il est fécondable 24 heures environ par un spermatozoïde, dans le 1er tiers de la trompe de Fallope. Il contient 23 chromosomes de la mère dont le déterminant sexuel féminin, X.
Dès l’entrée en contact du spermatozoïde avec la zone pellucide de l’ovule, l’acrosome du spermatozoïde se rompt, libérant un enzyme (ZP3) qui digère cette zone pellucide complexe recouvrant l’ovocyte. Le noyau du spermatozoïde se détache alors du flagelle et entre seul dans l’ovocyte 1 qui devient ovocyte 2. De même que lors d’un contact avec un spermatozoïde de race non humaine, afin de préserver l’ovule d’une double introduction de spermatozoïde (polyspermie), la membrane plasmique de l’ovocyte devient alors immédiatement infranchissable à un deuxième spermatozoïde.
Le noyau du spermatozoïde contient 23 chromosomes du père, dont le déterminant sexuel X ou Y selon le spermatozoïde concerné (Chromosomes féminins = XX, masculin = XY). L'ovocyte fécondé, appelé à ce moment ovocyte 2, continue d’évoluer durant sa descente dans la trompe de Fallope. Seize à dix-huit heures après l’ensemencement de l'ovule, les 2 noyaux cellulaires issus de l’ovule et du spermatozoïde, sont bien visibles dans la cellule avant de fusionner et devenir un.
Trente heures environ après la fécondation, le zygote (ovule fécondé) commence une division cellulaire (segmentation, clivage), selon une périodicité de 12 à 16 heures, sans que la taille extérieure de la zone pellucide (zone de peau) n’augmente. A l’intérieur de cette zone une réserve de vitelline (matière nutritive) est contenue entre deux membranes, ce qui assure la vie et l’énergie aux cellules jusqu’au raccordement sanguin de l’utérus. Ces cellules, appelées blastomères, deviennent donc de plus en plus petites.
A ce stade, les cellules (blastomères) sont encore de forme cylindrique. Elles continuent de se diviser au même rythme sans augmentation donc du volume total. Chacune d’elles conserve la totale polyvalence à générer quelque cellule d’organe spécifique que ce soit. Ce sont donc toutes des cellules totipotentes, c'est-à-dire du plus haut niveau de polyvalence parmi les cellules souches (pluripotentes, multipotentes ou unipotentes).
Jusqu'au stade de morula, les cellules embryonnaires sont totipotentes (cellules souche non spécialisées), c'est-à-dire que chaque cellule pourra donner n'importe quelle cellule spécialisée (peau, os, muscle, cerveau, placenta...), ainsi que les annexes embryonnaires de l'embryon comme le placenta, ce qui n'est plus possible par la suite aux cellules pluripotentes qui deviendront spécialisées dans l’évolution de l’embryon.
C’est à ce stade, par une trop rapide évolution des cellules, que peut se produire une division de la morula et donner naissance à de vrais jumeaux (ou jumeaux monozygote). Ils seront alors obligatoirement de la même sexualité, contrairement aux jumeaux nés de deux ovules différents.
A l'approche du 5ème jour après la fécondation, une autre transformation décisive survient. La morula subit une accumulation de liquide qui forme par confluence une cavité interne appelée blastocœle. L’ensemble est appelé blastula puis blastocyste au moment de l’implantation.
Au stade d'une dizaine de cellules, celles-ci entreront en compaction. Ce terme définit le passage de cellules d'une morphologie sphérique à une forme dite pavimenteuse grâce à laquelle, leurs surfaces de contact s'étendent. A ce stade de 8 à 16 cellules le zygote (ovule fécondé), prend alors le nom de morula (en raison de sa ressemblance à une mûre).
La cavité interne (blastocœle ou blastocele) va se dilater petit à petit et distendre la couche monocellulaire ainsi que la zone pellucide qui l'entoure encore (peau qui le protège, mais qui ne lui permettrait pas de s’implanter dans l’endomètre de l’utérus). Cette expansion du blastocyste (ensemble embryonnaire), constitué alors d’une centaine de cellules, se produit vers la fin du 5ème jour, jusqu'à rupture de la zone pellucide (1) : c'est l'éclosion blastocytaire qui permettra la nidation dans l’endomètre de l’utérus. Cet ensemble embryonnaire est formé par la couche monocellulaire de cellules trophoblastes (2) bordant la cavité interne qui sera le point de départ du placenta et par un amas de cellules embryoblastes (3) à partir desquelles de développera l’embryon puis le fœtus.
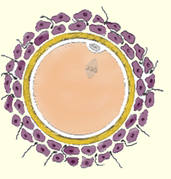
Représentation de la Zone Pellucide de l’ovocyte 1 et des cellules péri-ovocytaires de la granulosa lors de l’ovulation
La zone pellucide, que nous avions jusqu’alors assimiler à une quelconque pellicule protectrice, est en réalité beaucoup plus complexe, si nous la regardons en détail. La structure globale ci-dessous, met en évidence, la partie mécanique de protection et de sélection, à laquelle s’ajoutent cependant des variations hormonales, ainsi que les différents enzymes mis à contribution simultanément selon les étapes. Selon vos désirs, nous vous invitons volontiers à approfondir sur les sites scientifiques spécialisés, ce qui ajoute à la complexité de l’ensemble.
Période ovarienne de l’ovule
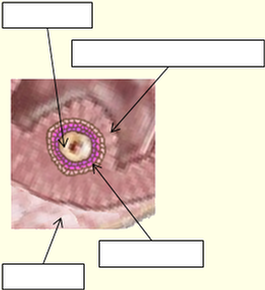
Follicule Antral (ou tertiaire)
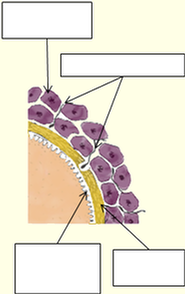
1) La zone pellucide qui a commencé de se créer au stade du follicule secondaire, s’entoure progressivement de cellules dite « granulaires » de forme relativement cubique. En continuant à se développer, elles vont former plusieurs couches cellulaires appelées granulosa. Dans les moments précédant l’ovulation, cette granulosa peu former plusieurs dizaines de couches autour de l’ovocyte. Une partie d’entre elles va donc se trouver entraînée au cours de l’ovulation, tandis que la partie restante va contribuer à produire les progestérones durant la dernière phase du cycle menstruel. En cas de fécondation, ces cellules associées au corps jaune de l’ovaire, vont servir à la production de ces progestérones jusqu’au moment où le placenta prendra le relais.
2) Les filaments retenant la première couche de la granulosa, se trouvent distendus au moment de l’expulsion de l’ovule hors du follicule et de l’ovaire. Seule la première couche de ces cellules reste solidaire de la zone pellucide, les couches supérieures formant un ensemble tissulaire appelé corona radiata, plus ou moins cohérent avec la première couche. Ces cellules granulaires sécrètent de la progestérone, ce qui est un appât pour les spermatozoïdes, au même titre que le liquide folliculaire.
3) Les grappes de cellules granulaires sont loin d’une régularité telle que représentée ci-contre, et comportent souvent plusieurs couches. Elles forment donc une sorte de barrière sélective, qui va permettre aux spermatozoïdes les plus forts d’atteindre l’ovule, mais aussi d’en limiter la fréquence d’approche, et servir de premier écran limitant la polyspermie.
4) L’ovule quant à lui n’étant pas source de progestérone, n’est donc pas un appât privilégié pour les spermatozoïdes. Dans l’avancée de ces spermatozoïdes, c’est un peu par hasard que certains d’entre eux vont donc venir en contacte de la zone pellucide. Sur les millions de spermatozoïdes émis lors d’un rapport sexuel masculin, seule une minorité d’entre eux (quelques dizaines maximum) vont ainsi plus ou moins progressivement risquer d’atteindre l’ovule et permettre la fécondation. Lors du contacte du spermatozoïde avec la zone pellucide de l’ovule, l’acrosome se rompt et libère des enzymes ZP3 qui digèrent l’épaisseur de la zone pellucide, avant de traverser l’espace péri-vitellin qui forme la réserve nutritive provisoire.
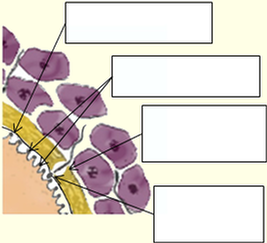
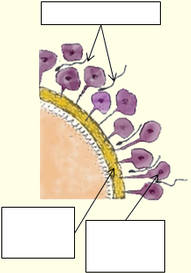
La zone pellucide se referme
Le noyau du spermatozoïde se détache
5) Le noyau du spermatozoïde se détache du flagelle et entre seul dans l’ovocyte, lors de la fécondation, ce qui provoque un éclatement des granules corticaux (diamètre 0,3 à 0,5 microns). L’éclatement de ces vésiculeuses granulés, situées sur toute la surface intérieure de l'ovocyte, libère leur contenu qui fusionne avec la membrane plasmique, ce qui est appelée réaction corticale pour former la membrane de fécondation, empêchant ainsi l'entrée de nouveaux spermatozoïdes, et donc la polyspermie. Pour certaines races non humaines, cette barrière contre la polyspermie est de nature électrique et non chimique. L’ovule change alors immédiatement de positivité lors de la fécondation, ce qui souligne bien la nature instantanée de cette fonction.
Les jours indiqués ci-dessous tiennent compte d'une fécondation immédiatement après ovulation, mais peuvent être décalés de 24 heures, ce qui correspond à la durée de vie de l'ovule avant fécondation.
Haut
Haut
Haut



